DOI:https://doi.org/10.46502/issn.2710-995X/2021.6.04
Herramienta para la obtención de Patrones de Conectividad Cerebral en Pacientes con Deterioro Cognitivo
Tool for obtaining Brain Connectivity Patterns in Patients with Cognitive Impairment
Resumen
Las Enfermedades Neurodegenerativas son patologías crónicas que producen discapacidad física, intelectual y social, dependencia y disminución de la calidad de vida de la persona afectada y sus familiares, tiene significativa repercusión laboral y económica. Estas se definen como un grupo de padecimientos de origen en muchos casos desconocido y que tienen como atributo común el curso progresivo de los síntomas derivados del daño estructural y funcional de diferentes zonas del sistema nervioso. La Demencia o Trastorno Cognitivo, ocurre cuando el cerebro se afecta por una enfermedad o afección específica que causa deterioro cognitivo. La secuencia en el espectro de la demencia puede terminar en la forma más grave de la enfermedad. El deterioro cognitivo en muchos casos transita desde el leve, referido a una disminución de la función cognitiva mayor de lo esperado por la edad y al nivel de educación del individuo, pero sin interferir con sus actividades cotidianas. El diagnóstico temprano de la enfermedad y la asunción de tratamientos y sobre todo estilos de vida saludables, puede evitar o enlentecer el tránsito hacia la gravedad de la enfermedad y por ende mejorar la calidad de vida. Con el objetivo de contribuir a este propósito se diseñó una herramienta de software denominada Neuromap que permite obtener patrones de conectividad a partir de señales de EEG y realizar una prueba de profundidad de la demencia. De esta forma se enriquecen los instrumentos con que cuentan en la actualidad los especialistas de las ciencias médicas para obtener un mejor y más eficaz diagnóstico. La herramienta fue desarrollada en Matlab, utilizando herramientas de estudio del EEG.
Palabras clave: Deterioro Cognitivo Leve, Deterioro cognitivo mayor, Enfermedades Neurodegenerativas, Enfermedad de Alzheimer, Patrón de conectividad cerebral.
Abstract
Neurodegenerative Diseases are chronic pathologies that produce physical, intellectual and social disabilities, dependency and decrease in the quality of life of affected person and their family with a significant labor and economic repercussion. These are defined as a group of diseases of unknown origin in many cases and which have as common attribute the progressive course of the symptoms derived from the structural and functional damage of different nervous system areas. Dementia or Cognitive Disorder occurs when the brain is affected by a specific disease or condition that causes cognitive decline. The sequence on the spectrum of dementia can end in its severe form. In many cases Cognitive impairment transits from mild, referring to a decrease in cognitive function greater than expected by the age and level of education of an individual, but without interfering with daily activities. The early diagnosis of the disease and the assumption of treatments and, above all, healthy lifestyles, can avoid or slow down the transit towards the severity of the disease and therefore, improve the quality of life. In order to contribute to this purpose, was designed a software tool named Neuromap to obtain connectivity patterns from EEG signals and perform a dementia depth test. In this way, was enriched the instruments currently available to medical specialists to obtain a better and more efficient diagnosis. The tool was developed with Matlab, using the EEG study tools.
Keywords: Alzheimer Disease, Brain connectivity pattern, Mild Cognitive impairment, Major Cognitive Impairment, Neurodegenerative Disease.
Introducción
Las Enfermedades Neurodegenerativas (EN) son padecimientos crónicos que tienden a generar discapacidad física, intelectual y social y, por tanto, dependencia y deterioro de la calidad de vida. Lo que va más allá de la afectación al paciente, alcanzando también a su familia. Estos padecimientos tienen una repercusión significativa en la actividad laboral y la economía por la atención familiar, social y sanitaria que precisan.
Las EN se definen como un grupo heterogéneo de enfermedades de causa aun no completamente esclarecida y que tienen como atributo común el curso progresivo de los síntomas de deterioro cognitivo, derivados del daño estructural y/o funcional del sistema nervioso central o periférico (Rodriguez Escobar et al, 2016).
Entre las EN, la demencia o Trastorno Cognitivo Mayor (TCM), ocurre cuando el cerebro se afecta por una enfermedad o afección específica que causa deterioro cognitivo. Al estudio de dichas enfermedades se le ha otorgado gran importancia debido a la tendencia al envejecimiento poblacional que se manifiesta a nivel global.
Se estima que en mundo existen 23,4 millones de personas con demencia (Capote Leyva et al, 2016), en las próximas décadas esta cifra aumentará ya que se aprecia un crecimiento exponencial de la enfermedad, duplicándose su incidencia cada 5 años. Prevalecen los enfermos a partir de los 65 años, aunque afecta también a personas jóvenes, y predomina el sexo femenino (Garre-Olmo, 2018).
El espectro de la demencia se puede apreciar como una secuencia de problemas cognitivos que comienza con deterioro cognitivo leve (DCL) y termina con demencia severa o Trastorno Cognitivo Mayor (TCM). El DCL se establece como una disminución de la función cognitiva mayor de lo esperado, con respecto a la edad y al nivel de educación de un individuo, pero que no interfiere con sus actividades diarias (Arriola Manchola et al., 2017). Clínicamente, es la etapa temprana de transición entre la cognición normal y el TCM.
En Cuba la demencia y la enfermedad de Alzheimer durante los años 2017 y 2018 causaron la muerte de 5 037 y 5 477 personas respectivamente, ubicándose como la sexta causa de muerte en el país (ANUARIO ESTADÍSTICO DE SALUD 2018, 2019). La prevalencia de la demencia en la población mayor de 65 años se sitúa entre 6,4 y 10,8 %, constituyendo la de tipo Alzheimer su causa más frecuente. Se estima que en los próximos 15 años, la población mayor de 80 años ascenderá a casi medio millón de personas, de la cual alrededor del 40 % padecerá la enfermedad de Alzheimer, de manera que si no se encuentra una solución en los próximos años, en el año 2020 se duplicará la cifra existente hoy en día que asciende a 100 000, y que con este ritmo llegara a 350 000 para el año 2040 (Capote Leyva et al, 2016).
El procesamiento de los registros de Electroencefalografía (EEG) se vislumbra como herramienta efectiva para el diagnóstico desde etapas tempranas del trastorno cognitivo.
El método de obtención, a partir de estos registros, de patrones de conectividad cerebral en estado de reposo cuya obtención constituye antecedente de la presente investigación (Garcia Leyva, 2018), permite diferenciar de manera efectiva los trastornos cognitivos y sus etapas, siendo una alternativa para elevar la eficacia y oportunidad de los diagnósticos, beneficiando así a la salud del paciente y por consiguiente la situación familiar.
Hasta el momento este método no contaba con una herramienta computacional que facilitara su uso por especialistas de salud e investigadores en la temática, de ahí que se abordó el siguiente problema científico: No se cuenta con una herramienta de software capaz de analizar la conectividad cerebral de pacientes en estado de reposo con trastornos cognitivos.
Teniendo como objeto de la investigación: Diseño de software para diagnóstico de enfermedades neurodegenerativas en el que se precisa como campo de acción: Diseño de software para el diagnóstico de trastornos cognitivos.
Para dar solución al problema identificado se propuso como objetivo general:
Diseñar software para el diagnóstico de trastornos cognitivos en estado de reposo a partir del procesamiento digital de señales de EEG y la obtención de los patrones de conectividad cerebral en pacientes con trastornos cognitivos.
Se definieron como objetivos específicos:
•Definir los criterios de diagnóstico que se ofrecerán al especialista
•Obtener los patrones de conectividad a partir del procesamiento de la señal de EEG.
•Desarrollar la herramienta de software que permita obtener los patrones de conectividad cerebral y ofrecer elementos de apoyo al diagnóstico.
El problema señalado, así como la precisión del objetivo general en esta investigación responden a la hipótesis: Es posible diseñar un software para obtener patrones de conectividad cerebral mediante el método de mapeo cerebral que auxilie al especialista en el diagnóstico de trastornos cognitivos.
Marco Teórico
La conectividad cerebral es un término referido a tres aspectos fundamentales: el patrón de enlaces anatómicos (Conectividad Anatómica), el patrón de dependencias estadísticas (Conectividad Funcional), o a el patrón de interacciones causales entre unidades distintas dentro del sistema nervioso (Conectividad Efectiva), estas unidades se corresponden con neuronas individuales, poblaciones neuronales, o regiones anatómicamente segregadas del cerebro (Sporns, 2007).
Los patrones de conectividad están formados por los enlaces que se establecen por medio de la sinapsis o por los enlaces a través de haces de fibra, o representando relaciones estadísticas o causales que pueden ser medidas a través de correlación, coherencia o flujo de información. La actividad neuronal y, por extensión, los códigos neuronales, están limitados por la conectividad, por lo que esta es crucial para dilucidar cómo las neuronas y las redes neuronales procesan la información(Horwitz, 2003).
La conectividad anatómica está referida a una red de conexiones físicas o estructurales (sinápticas) que unen conjuntos de neuronas o elementos neuronales, así como sus atributos biofísicos estructurales asociados. El patrón físico de las conexiones anatómicas es relativamente estable en escalas de tiempo pequeñas (segundos a minutos). En escalas de tiempo más largas (de horas a días), los patrones de conectividad estructural probablemente están sujetos a los cambios morfológicos que ocurren en el cerebro. Es importante señalar que solo los estudios de rastreo invasivo actualmente son capaces de demostrar las conexiones axonales directas (Garcia Leyva, 2018).
La conectividad funcional, es fundamentalmente un concepto estadístico, en general captura las desviaciones de la independencia estadística entre las unidades neuronales (distribuidas y espacialmente remotas), o sea, se verifica que la actividad de dos estructuras cerebrales está relacionada, de lo contrario, si la actividad de una estructura no depende de la actividad de la otra, entonces se dice que tienen independencia estadística. Esta puede estimarse midiendo la correlación o la covarianza, la coherencia espectral o la sincronización de fase. La conectividad funcional es altamente dependiente del tiempo y a menudo se calcula entre todos los elementos de un sistema, independientemente de si estos elementos están conectados por enlaces estructurales directos o no (Sporns, 2007).
La conectividad efectiva se puede interpretar como la unión de la conectividad estructural y la funcional, ya que describe las redes de efectos direccionales de un elemento neuronal sobre otro. Los principios aplicados para su evaluación son los efectos causales que pueden inferirse a través de perturbaciones sistemáticas del sistema o a través del análisis de series de temporales, dado que las causas deben preceder a los efectos en el tiempo (McIntosh & Mišić, 2013).
Formalmente, los patrones de conectividad cerebral se pueden representar en forma de grafos, los cuales pueden ser ponderados, con pesos que representen diversos parámetros, o representarlos de forma binaria, con elementos binarios (nodos y aristas) que indican la presencia o ausencia de una conexión (Garcia Leyva, 2018).
La relación entre la conectividad anatómica, funcional y efectiva en la corteza representa un desafío significativo para la neurociencia teórica actual. Dos principios potenciales que vinculan estos diferentes modos de conectividad cerebral son la segregación y la integración (Tononi, Sporns, & Edelman, 1994). La segregación se refiere a la existencia de neuronas y áreas cerebrales especializadas, organizadas en distintas poblaciones neuronales y agrupadas para formar áreas corticales. El principio complementario, la integración, da lugar a la activación coordinada de poblaciones neuronales distribuidas, lo que permite el surgimiento de estados cognitivos y conductuales coherentes. La interacción de la segregación y la integración en redes cerebrales genera información que a la vez está muy diversificada y altamente integrada, creando así patrones de alta complejidad (Shine, Aburn, Breakspear, & Poldrack, 2018).
Análisis de conectividad: métodos, modelos y técnicas
Los métodos computacionales para el procesamiento del EEG analizan la conectividad cerebral en dos dominios o espacios diferentes, uno de ellos es el espacio “Electrodos” (Scalp), el cual genera discrepancias y polémica en la comunidad científica, relacionadas con el proceso de inferir las interacciones de las diferentes estructuras cerebrales debido a los efectos que provoca la conducción del volumen cerebral, por lo que se plantea que reconstruir las fuentes de la actividad cerebral a partir de los datos registrados, o sea, analizar la conectividad en el domino fuente (Source) es la forma más precisa de inferir las interacciones entre diferentes estructuras (Garcia Leyva, 2018).
Evidencias básicas de la fisiología cortical permiten establecer algunas predicciones sobre las fuentes corticales de las señales de EEG. La corteza posee una densidad extremadamente alta de conexiones locales (Shine et al., 2018) las cuales están eléctricamente alejadas del cuero cabelludo donde se registra el EEG, por lo que es más probable que la fuente de los registros sean regiones compactas de corteza cuya actividad eléctrica local, a escala neuronal, está similarmente orientada por la geometría cortical y parcialmente sincronizada en el orden de cm2, sin embargo producen señales que pueden alcanzar el cuero cabelludo por medio de la conducción del volumen cerebral. Todo lo expuesto permite la aproximación a un modelo de fuente de actividad cerebral que es compatible con la fisiología cortical (Jones, 2000; Stettler, 2002).
Separación de fuentes de actividad cerebral mediante ICA
La fase de preprocesado de una señal EEG no garantiza la eliminación total de todos los artefactos. Por ello, se debe acudir a técnicas de reducción de dimensionalidad. En este tipo de técnicas la información redundante o innecesaria de cada canal es eliminada, facilitando así la búsqueda de información relevante. El análisis de componentes independientes (ICA) es una herramienta útil para descomponer señales de EEG en patrones de actividad con una máxima independencia, los cuales son compatibles con la actividad en una sola área cortical activa (Delorme et al., 2011; Makeig, Debener, Onton, & Delorme, 2004). Existen bases biológicas que sustentan esta afirmación, las cuales plantean que en circunstancias favorables, ICA debe separar las señales que surgen de las actividades de campo locales, en áreas de fuentes corticales compactas y físicamente distintas, basadas en el hecho de que las conexiones laterales de corto alcance (<100 µm) entre las neuronas corticales son mucho más densas que las conexiones de mayor alcance (Stepanyants, Martinez, Ferecskó, & Kisvárday, 2009; Stettler, 2002), mientras que las redes de células gliales e inhibitorias no tienen procesos de largo alcance. Además, las conexiones tálamo-corticales que también desempeñan un papel importante en la dinámica del campo son predominantemente radiales (Garcia Leyva, 2018).
Este método ofrece una teoría de la dinámica cerebral mediante la separación de la actividad independiente en cada registro de EEG. Este proceso se realiza no por medio de un filtrado espacial directo para la actividad generada en un grupo de localizaciones corticales predefinidas, sino mediante el uso de la información contenida en los datos en sí mismos, para separar porciones de datos registrados en el cuero cabelludo a partir de cada área activa o fuente de actividad cortical o artefactos, basándose en la asunción estadística y fisiológicamente probable, de que en el tiempo, estas actividades deben ser o estar cerca, de la independencia la una de la otra (Wessel, 2018).
La principal ventaja de esta teoría es que la actividad localmente coherente, constituyente de una sola fuente de EEG, puede ser agrupada en una sola componente independiente que incluye su proyección a todos los canales, mientras que la actividad de las fuentes de EEG no relacionadas, puede ser descartada de esta componente independiente y aislada a otra componente independiente. De esta forma, bajo circunstancias favorables, ICA puede transformar datos registrados con una alta densidad de electrodos, en un grupo de fuentes corticales y artefactos por separado (Garcia Leyva, 2018).
Cuando se aplica ICA al procesamiento de datos de EEG es necesario establecer ciertas consideraciones relacionados con la fisiología del fenómeno en cuestión. Una suposición general y fisiológicamente posible, presente en la mayoría de los análisis relacionados con el EEG, es que la mayoría de los potenciales de campo lejano, (far-field potentials) detectados en el cuero cabelludo, no fueron generados en el cuero cabelludo en sí mismo, pero sí dentro de su dominio espacial, o dígase en las áreas donde las neuronas corticales piramidales están similarmente orientadas. Un área cortical dada solo puede producir un potencial de campo lejano, y de este modo convertirse en una fuente de EEG, si los potenciales locales de las células piramidales que la rodean están parcialmente sincronizados (no es necesario que lo estén completamente) (Pesaran et al., 2018).
En la aplicación de ICA en este trabajo se asume que la actividad registrada en el cuero cabelludo es producida por fuentes corticales de EEG, la cual es proyectada casi instantáneamente al cuero cabelludo por medio del efecto de conducción del volumen cerebral. Esto significa que las señales registradas en el cuero cabelludo, son la mezcla de todos los potenciales originados en las áreas corticales cercanas, más las fuentes producidas por artefactos(Garcia Leyva, 2018).
Localización de fuentes de actividad cerebral
La comprensión del proceso de localización de fuentes de actividad cerebral requiere, primeramente, comprender los procesos que suceden a nivel biológico en el cerebro. El bloque de construcción principal del sistema nervioso humano es la neurona, y como en todas las demás células del organismo humano, el cuerpo celular de las neuronas contiene la información genética, así como una infraestructura para transportar fuera los productos del metabolismo básico de la célula.
Lo que separa a las neuronas de las otras células típicas es la presencia de dendritas y de un axón. Las dendritas son el lugar donde las neuronas integran la energía proveniente de otras células, la cual puede ser excitatoria o inhibitoria. Cuando un cierto umbral de energía es sobrepasado, es en el axón donde el potencial de acción se propaga, lo que significa que un neurotransmisor específico es liberado hasta el final de este a través de una estructura denominado sinapsis. Los neurotransmisores liberados a partir de la sinapsis están ligados a los receptores presentes en las dendritas de alguna otra neurona, la cual refleja una potencial post-sinapsis (PSP).
En la actualidad se conoce que las diferencias de potencial eléctrico en el cuero cabelludo son causadas por el PSP y están determinadas por la sumatoria espacial de la densidad de corriente proveniente de los potenciales post-sinapsis que ocurren sincrónicamente en un grupo de neuronas (Pascual-Marqui, 2009). Aunque todas las neuronas contribuyen a las señales de EEG, estas son primariamente generadas por neuronas en la superficie cortical (neuronas piramidales corticales) que están orientadas perpendicularmente a la superficie. Por otra parte, una sola neurona activa no es suficiente para generar una señal de EEG medible, solo cuando un grupo de neuronas piramidales corticales están dispuestas en paralelo y sincronizadas se puede, a través de la electroencefalografía, detectar una señal.
Estimadores de Conectividad Cerebral
En la actualidad se han desarrollado diferentes estimadores de conectividad cerebral (Sakkalis, 2011) y para su estudio en esta tesis se han dividido en estimadores de conectividad funcional y de conectividad efectiva.
Estimadores de Conectividad Funcional
La conectividad funcional del cerebro se puede estimar de varias maneras, por ejemplo, a través de correlaciones cruzadas en el dominio del tiempo o la frecuencia, información mutua o coherencia espectral. A continuación, se presenta una posible taxonomía de las métricas comúnmente utilizadas para estimar la conectividad funcional, figura 1.1. Una subdivisión se puede hacer en base a la métrica que cuantifica la dirección de la interacción. Las métricas de conectividad funcional no dirigida buscan capturar alguna forma de interdependencia entre las señales, sin referencia a la dirección de la influencia, por el contrario, las métricas de conectividad funcional dirigida tratan de establecer una causalidad estadística a partir de los datos, lo cual está basado en el precepto de que las causas preceden a sus efectos, y en el caso de la causalidad de Granger y la transferencia de entropía, las causas pueden en alguna manera predecir sus efectos.
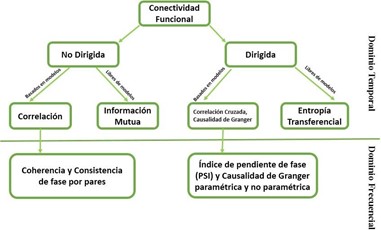
Fig. 1.1. Métricas comúnmente utilizadas para estimar la conectividad funcional. Tomado de (Bastos & Schoffelen, 2016).
Estimadores de Conectividad Efectiva
La conectividad efectiva es una progresión lógica desde la conectividad funcional y se refiere a la influencia directa de un elemento neuronal en otro. La principal distinción entre conectividad funcional y efectiva radica en los niveles de inferencia que permiten. La conectividad funcional, estimada como dependencias estadísticas entre elementos neuronales, no permite la inferencia sobre la direccionalidad de la influencia, mientras que la conectividad efectiva se estima mediante la formulación de un modelo causal explícito de cómo los elementos neuronales interactúan entre sí, lo que permite la inferencia sobre las influencias direccionales.
Esto se ilustra en la figura 1. 2, en la que las dos redes anatómicas difieren en un aspecto: la presencia de una proyección directa desde el nodo A al nodo B (McIntosh & Korostil, 2008). Los nodos A y B reciben entradas comunes y, como resultado, mostrarán conectividad funcional en ambas redes. Por lo tanto, las dos redes no podrían diferenciarse en términos de conectividad funcional, pero sí en términos de conectividad efectiva. El propósito de la conectividad efectiva es modelar explícitamente la direccionalidad en la red (Vecchio et al., 2017)

Fig. 1.2. Dos configuraciones de redes diferentes, tomado de (Mišić & McIntosh, 2015)
Las redes tienen topologías similares, salvo una proyección de A hacia B, que está ausente en la red 1 y está presente en la red 2. Como A y B reciben entradas comunes de C, la conectividad funcional no podría distinguir entre las dos configuraciones. La conectividad efectiva, que modela explícitamente la direccionalidad de la influencia, podría determinar si existe alguna influencia residual de A y B, una vez que se tengan en cuenta los efectos intermedios de C.
Entre las técnicas empleadas para estimar la conectividad efectiva están los métodos confirmatorios, en el sentido de que se formula y prueba un modelo explícito, de las interacciones de los elementos, para ver si se ajusta a los datos observados. Estos métodos son el modelado de ecuaciones estructurales (SEM) y el modelado causal dinámico (DCM). Las otras técnicas son las basadas en la causalidad de Granger, empleadas para estimar la influencia causal entre elementos. Estas se aplican, generalmente, de manera exploratoria en cualquier par de elementos neurales (Garcia Leyva, 2018).
Resultados y discusión
Como resultado de la investigación se desarrolló el software “NeuroMap” capaz de realizar el análisis de conectividad cerebral para determinar la existencia de signos de deterioro cognitivo.
Descripción General del Sistema
La interfaz gráfica de usuario desarrollada “NeuroMap” se utiliza para el análisis de la señal de EEG y la presentación de los resultados obtenidos, está diseñada para la investigación y el diagnóstico de DCL mediante el método de mapeo cerebral, con el fin de dar al usuario un criterio adicional para emitir un diagnóstico. Posee opciones para la visualización de gráficos animados y estáticos de las fuentes de Conectividad, de la señal de EEG y sus respectivos episodios. La misma integra además una prueba para determinar la profundidad de la demencia (Test Mini-mental). La herramienta cuenta con un flujo de procesos como se muestra en la figura 1.3.
Se muestran la densidad espectral de potencia, el resultado del análisis de componentes independientes las fuentes de conectividad en forma de imágenes que reflejan la topografía craneal y la posibilidad de eliminar los artefactos causados por diversas causas obteniendo así resultados más favorables. Los resultados del análisis de conectividad se presentan de forma animada, figura 1.5,1.6,1.7 con la posibilidad de salvar las imágenes en el formato deseado. La herramienta ofrece la posibilidad de guardar estos resultados en un reporte en formato Word para que pueda ser estudiado por el especialista.

Fig.1.3. Diagrama de Flujo de herramienta Neuromap.
Análisis de Conectividad La corteza posee una densidad extremadamente alta de conexiones locales las cuales están eléctricamente alejadas del cuero cabelludo donde se registra el EEG, por lo que es más probable que la fuente de los registros sean regiones compactas de corteza cuya actividad eléctrica local, a escala neuronal, está similarmente orientada por la geometría cortical y parcialmente sincronizada en el orden de cm2, sin embargo producen señales que pueden alcanzar el cuero cabelludo por medio de la conducción del volumen cerebral. Siguiendo esta premisa en esta ventana con diagrama de flujo de procesos mostrado en la figura 1.4 se realiza el análisis de conectividad a partir del registro de EEG introducido por el usuario en formato (.mat).

Fig. 1.4. Diagrama de flujo de Ventana Análisis de Conectividad.

Fig. 1.5. Conectividad cerebral entre fuentes en un paciente sano.

Fig. 1.6. Conectividad cerebral entre fuentes en un paciente con EA.
Análisis de conectividad cerebral en pacientes con Deterioro Cognitivo Leve
La conectividad cerebral fue analizada en todos los grupos estudiados con el objetivo de identificar patrones de conexión característicos en dichos grupos. Nuestro trabajo apoya la opinión de que las alteraciones cognitivas asociadas a la EA pueden deberse no solo a la pérdida de neuronas, sino también a alteraciones en la coordinación temporal en la distribución de la actividad neurona (Uhlhaas & Singer, 2006).
Un enfoque diferente es analizar el flujo de información electrofisiológica en el dominio fuente. Con el objetivo de abordar los resultados obtenidos en nuestra investigación se seleccionaron tres pacientes (uno de cada grupo), lo cuales representan las características encontradas en cada grupo estudiado.
La figura 1.5 nuestra la vista sagital de un modelo de cerebro promedio (estándar NMI) en la cual se han representado las fuentes de actividad cerebral estimadas en un sujeto del grupo de pacientes supuestos sanos.
En la figura anterior se puede apreciar que en el dominio fuente se obtuvieron resultados consecuentes con los que se obtuvieron en el dominio sensor o electrodo, pues en esta figura se puede observar que la fuente número 4, en el intervalo de tiempo representado (0,4 segundos), es la fuente que está predominantemente activa y se encuentra localizada en la región posterior.
Las métricas relacionadas con la teoría de grafos, definidas en (Bullmore & Sporns, 2009; Seth, 2005), y que fueron aplicadas en esta investigación, muestran dos componentes, los nodos y las aristas (edges), que en este caso representan los enlaces entre las fuentes. La información proporcionada por el análisis mediante teoría de grafos y que está contenida en la figura anterior es: flujo de salida (outflow), representado mediante el tamaño de los nodos, y flujo causal (causalflow), representado mediante el color de los nodos.
El flujo de salida es la suma de la fuerza o fortaleza de la conectividad sobre los enlaces (aristas) en dirección saliente del nodo, y el flujo causal es el flujo de salida menos el flujo de entrada (inflow), el cual está definido por la suma de la fuerza o fortaleza de la conectividad sobre los enlaces (aristas) en dirección entrante del nodo.
Los valores obtenidos al aplicar el estimador de conectividad seleccionado, la dDTF, están codificados de igual manera mediante el grosor de las aristas (los enlaces que unen los nodos) y el color de estas.
El nodo 4 (fuente número 4) en el instante de tiempo analizado, presenta el valor máximo en cuanto a flujo de salida y el valor máximo en cuanto a flujo causal pero además tiene el máximo valor de conectividad en dirección a las otras fuentes. Estos resultados son consecuentes con los que se encontraron en el dominio electrodos para los pacientes del grupo de sujetos supuestos sanos, y reflejan los patrones de conectividad que fueron establecidos para este grupo, donde se pudo observar que la actividad cerebral para el estado de reposo con cojos cerrados que generaba en la región posterior del cerebro principalmente y luego se producía un flujo de información causal hacia el resto de las regiones cerebrales sobre todo hacia la región anterior.
Estos resultados están acordes con otros estudios relacionados con el estado cerebral analizado donde se han obtenido mediante EEG y validado mediante fMRI, las fuentes de la actividad cerebral y el flujo de información electrofisiológica entre estructuras cerebrales (Knyazev, Slobodskoj-Plusnin, Bocharov, & Pylkova, 2011; Mahjoory et al., 2017).
La figura 1.6 muestra la vista sagital en el modelo de cerebro promedio (estándar NMI), en el cual se han representado las fuentes de actividad cerebral estimadas en un sujeto del grupo de pacientes con DCL.
Al analizar las métricas obtenidas mediante la teoría de grafos podemos ver el nodo 4 (fuente número 4), localizado también en la región posterior del cerebro, en el instante de tiempo analizado, presenta un valor máximo en cuanto a flujo de salida y un valor máximo en cuanto a flujo causal pero además tiene el máximo valor de conectividad en dirección a una de las otras fuentes localizadas. Estas diferencias de conectividad, con relación a los pacientes sanos, no son considerables, pues aún en este estadio inicial de la enfermedad sus características no han manifestadas de forma precisa.
Los resultados antes expuestos evidencian que en este grupo de estudio no se encontraron diferencias en cuanto al flujo de información causal, de la región posterior a la región anterior en relación con el grupo de pacientes sanos en el dominio fuente.
La figura 1.7 muestra la vista sagital en el modelo de cerebro promedio (estándar NMI), en el cual se han representado las fuentes de actividad cerebral estimadas en un sujeto del grupo de pacientes con EA. En este grupo de pacientes fue donde principalmente se encontraron fuentes de actividad cerebral en regiones atípicas a las encontradas en los pacientes sanos para el estado cerebral analizado en esta investigación, por lo que la conectividad en el domino fuente también refleja esas características anormales.
Al analizar las métricas obtenidas mediante la teoría de grafos podemos ver que en este caso el nodo 4 está localizado en la región temporal, y a pesar de que su tamaño e intensidad de color sugieren un valor elevado en su flujo de salida y en su flujo causal, se hace notar que la escala de medición difiere a los casos anteriores pues fue necesario disminuirla para que estas magnitudes pudieran ser apreciadas. Al realizar un análisis en detalle se puede apreciar que tanto el flujo de salida y el flujo causal, así como los valores de conectividad en las fuentes localizadas en la región posterior disminuyeron en relación con el grupo de pacientes sanos, solo observa un flujo de información desde la fuente 4 en la región temporal derecha hacia la región posterior, lo cual no es típico del estado cerebral analizado.
En estos pacientes no se aprecia el flujo causal de información desde la región posterior hacia la región anterior, que fue encontrado en los sujetos del grupo supuesto sano. Este fenómeno pudiera deberse a procesos relacionados con la atrofia cortical posterior (PCA) la cual ocurre cuando están afectadas áreas en la región posterior del cerebro, específicamente los lóbulos parietal y occipital (Lopez, 2014). Además, es conocido que la EA provoca la acumulación fragmentos de proteínas beta-amiloides (llamados placas beta- amiloides) fuera de las neuronas y la acumulación de una forma anormal de la proteína tau, dentro de las neuronas son dos de varios cambios cerebrales asociados con la enfermedad de Alzheimer.
Además, se plantea la hipótesis de que las placas beta amiloideas contribuyen a la muerte celular al interferir con la comunicación de neurona a neurona en las sinapsis, mientras que forma anormal de la proteína tau, bloquea el transporte de nutrientes y otras moléculas esenciales dentro de las neuronas (Alzheimer's & Dementia, 2017).
Otro de los aspectos para tener en cuenta es que los cerebros de las personas con EA avanzada muestran inflamación, reducción dramática de la pérdida de células y restos diseminados de neuronas muertas o moribundas.
Los resultados presentados en este trabajo sugieren la posibilidad de aplicar los métodos y metodologías estudiadas, sensibles a las características de la enfermedad, para la clasificación de pacientes con trastornos cognitivos y el desarrollo de biomarcadores predictivos del riesgo de evolucionar de una etapa inicial de la enfermedad a un estadio avanzado.
Conclusiones
Con el presente trabajo se dio cumplimiento a los objetivos propuestos:
1. Se creó la herramienta de software NeuroMap que permite obtener los patrones de conectividad cerebral mediante el método de mapeo cerebral que auxiliará al especialista en el diagnóstico de trastornos cognitivos.
2. La herramienta NeuroMap desarrollada incorpora elementos de apoyo al diagnóstico clínico con la incorporación de la prueba Mini-mental y el resultado del estudio de conectividad cerebral bajo los criterios previamente identificados como parte de la investigación.
3. A partir de los resultados obtenidos la herramienta puede ser de ayuda para diagnosticar casos de deterioro cognitivo leve ayudando así a reducir los posibles efectos de un diagnóstico tardío.
4. Por su diseño flexible la herramienta permite incorporar nuevas pruebas y métodos de procesamiento, en tanto avance la investigación.